.3. Meiosi, il processo base della riproduzione sessuata3.1 Evoluzione delle pianteEsaminando la diversità delle piante moderne, bisogna considerare che il passato rappresenta la chiave di lettura del presente. In questo senso, la storia delle piante è strettamente correlata allo sviluppo di adattamenti ai diversi habitat terrestri. In base ai reperti fossili, si possono distinguere quattro periodi principali che hanno caratterizzato l'evoluzione delle piante attuali. Ciascuno di questi periodi è contraddistinto dall'evoluzione di particolari strutture, la cui comparsa ha permesso un progressivo adattamento alla vita terrestre. Nel corso della storia ogni gruppo di piante che si è man mano evoluto ha sviluppato metodi di riproduzione sempre più adatti e complessi, fino alla riproduzione sessuata, che è il solo metodo che descriveremo in dettaglio. In questo albero filogenetico, i rappresentanti attuali delle principali linee evolutive delle piante sono illustrati in alto.

Il primo periodo evolutivo vide l'origine di piante da antenati acquatici, ossia dalle alghe verdi appartenenti al gruppo delle
carofite. Il primo adattamento alla vita terrestre fu lo sviluppo di particolari strutture chiamate
gametangi, destinate alla protezione dei
gameti e dell'
embrione in via di sviluppo. Ciò ha permesso ai vegetali noti come
briofite (i muschi) di diversificarsi dalle prime vere piante. Anche il tessuto vascolare si è evoluto relativamente presto. Tuttavia la maggior parte delle briofite ne è privo e viene pertanto classificata tra le piante non vascolari.
Il secondo periodo importante nell'evoluzione delle piante fu caratterizzato dalla diversificazione delle
piante vascolari (cioè provviste di tessuto specializzato per la conduzione dell'acqua e dei minerali). Le prime piante vascolari mancavano di
semi e i loro rappresentanti attuali sono le
felci e alcuni altri gruppi.
Il terzo periodo fondamentale iniziò con l'
origine dei semi. La produzione di semi permise la colonizzazione di ampie regioni di terraferma, poiché consentì una maggiore protezione dell'embrione dall'essiccamento e da altri fattori ambientali avversi. Un
seme è costituito dall'embrione racchiuso, insieme a sostanze di riserva, all'interno di un rivestimento protettivo. I semi delle prime piante a seme non erano tuttavia avvolti da ulteriori strutture specializzate. Si trattò di piante che dettero origine a molti gruppi di
gimnosperme (dal greco gymnos = nudo, e sperma = seme). Attualmente le gimnosperme più diffuse e diversificate sono le conifere.
Il quarto periodo che caratterizzò la storia evolutiva delle piante fu l'origine delle piante a fiore, o angiosperme (dal greco angion = vaso, e sperma = seme). A differenza dei semi nudi delle gimnosperme, il fiore è una complessa struttura riproduttiva contenente semi all'interno di camere protettive, detti
ovari. La grande maggioranza delle piante attuali sono angiosperme.
3.2 Le AngiospermeCome si può vedere dall'immagine precedente, le angiosperme, ovvero le piante con fiori sono state le ultime a svilupparsi e a colonizzare il nostro pianeta. Esistono circa 250.000 specie di angiosperme rispetto alle 700 specie di conifere e di altre gimnosperme. Il successo evolutivo delle angiosperme è quindi evidente, e dovuto ad alcuni adattamenti unici di queste piante. Il perfezionamento del tessuto vascolare, ad esempio, ha permesso di aumentare l'efficacia del trasporto idrico dalle radici alle foglie. Tuttavia, l'adattamento alla vita terrestre che ha maggiormente contribuito al successo di questo gruppo è stato lo sviluppo del
fiore.
3.3 - 3.4 Impollinazione e riproduzione delle orchideePer quanto riguarda la riproduzione sessuata e asessuata delle orchidee, la descrizione dell'apparato riproduttore (fiore, frutto e semi) e l'impollinazione si rimanda alle pagine 16-26 del libro Orchidee d'Italia, del G.I.R.O.S.
Per quanto riguarda l'impollinazione del genere
Orchis, si ricorda questo precedente approfondimento:
qui.
3.5 MeiosiA questo punto è importante avere ben chiaro la differenza tra cromosomi omologhi, cromatidi fratelli e cos'è la diploidia:
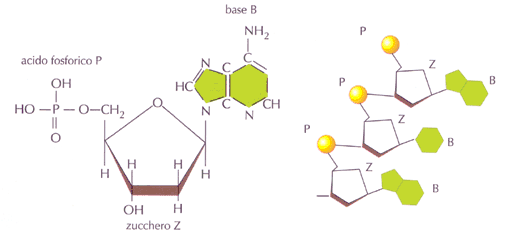
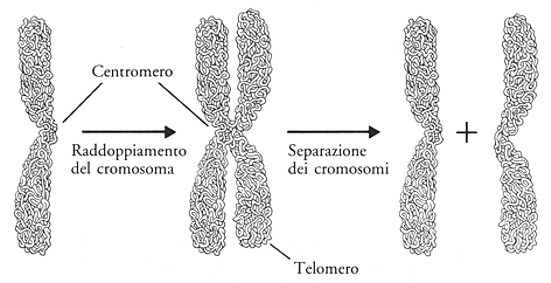
Come abbiamo già visto i
cromatidi fratelli si generano durante la mitosi, e sono i due "bastoncelli" di cui sono costituiti i cromosomi, si originano a seguito della duplicazione del DNA e pertanto contengono due molecole identiche di DNA.
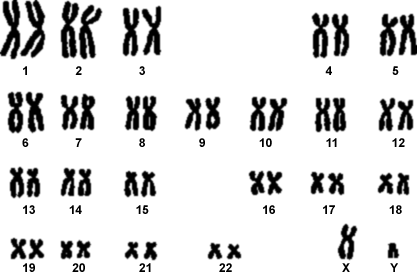
I cromosomi omologhi sono invece le coppie di cromosomi uguali presenti in una cellula diploide (2n). Nelle cellule umane tutti i cromosomi sono presenti in doppio, i due cromosomi omologhi, ereditati uno dal padre e uno dalla madre.
Il fatto di avere due corredi cromosomici, uno proveniente da ciascuno dei due genitori, costituisce un fattore chiave del ciclo vitale di tutte le specie caratterizzate da riproduzione sessuata, uomo ed orchidee comprese. Nel ciclo vitale la deposizione di polline su un altro fiore permette ai granuli pollinici (
gametofiti maschili) di unirsi alla cellula uovo presente nel sacco embrionale (
gametofito femminile) all'interno dell'ovario. Tale processo noto come fecondzione dà origine alla formazione della
cellula uovo fecondata diploide, o
zigote. Lo
zigote, possiede due corredi cromosomici omologhi, uno proveniente da ciascun genitore. Il ciclo vitale sessuato prevede l'alternanza tra stadi diploidi e aploidi, e la produzione dei
gameti aploidi, grazie alla meiosi, evita che a ogni generazione il numero di cromosomi raddoppi. A ogni generazione, il raddoppio del numero cromosomico che si avrebbe in seguito alla fecondazione, viene precedentemente compensato dal suo dimezzamento durante la meiosi.
Es. Nella
Orchis purpurea, il numero di cromosomi contenuti in una qualsiasi cellula della pianta diploide è 2n= 42, ad eccezione delle cellule destinate alla riproduzione (gametofiti maschili e femminali) che sono aploidi n= 21. In questo modo lo zigote che si forma dalla fecondazione (21 + 21) sarà nuovamente diploide e tutte le cellule della pianta che nascerà saranno diploidi (2n = 42). Se non avvenisse la meiosi, che dimezza il corredo genetico delle cellule destinate alla riproduzione la specie (gametofiti), ad ogni fecondazione la specie raddoppierebbe il corredo genetico (21 + 21... 42 + 42... 84 + 84...).
Vediamo ora un po' più nel dettaglio il processo della meiosi.
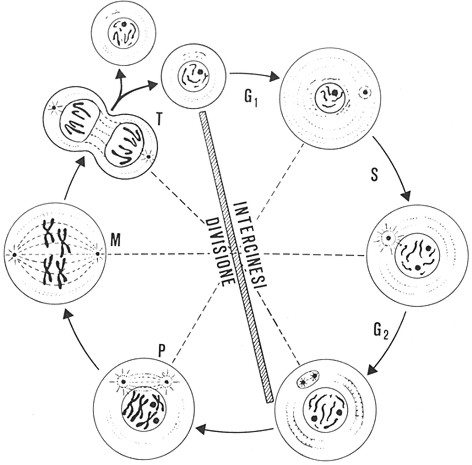
La
meiosi, ovvero il processo che produce gameti aploidi a partire da cellule diploidi è, per certi versi, simile alla mitosi, ma caratterizzato da due aspetti peculiari:
Il primo riguarda il dimezzamento del numero cromosomico. Durante la meiosi, una cellula cha ha precedentemente duplicato la propria cromatina subisce due divisioni consecutive, definite rispettivamente
meiosi 1 e
meiosi 2. Poiché queste due divisioni sono precedute da una sola duplicazione del materiale genetico (ved. replicazione del DNA, par. 2.5) ognuna delle quattro cellule figlie prodotte possiede soltanto metà dei cromosomi della cellula originale, ovvero un unico corredo cromosomico aploide. In realtà, il numero di cromosomi è aploide già al termine della meiosi 1, ma a tale stadio ogni cromosoma è ancora costituito da due cromatidi fratelli.
Il secondo aspetto caratteristico della meiosi riguarda lo scambio di materiale genetico, ovvero di frammenti di cromosomi, tra i due cromosomi omologhi di ogni coppia. Tale scambio, definito come
crossing over, ha luogo durante la profase della prima divisione meiotica, come vedremo a breve.
Si tenga ancora una volta ben presente la differenza tra cromosomi omologhi e cromatidi fratelli: i due cromosomi di una coppia di omologhi (2n) sono cromosomi interi ereditati da genitori differenti. I due cromosomi di una certa coppia (cromosomi omologhi) portano la medesima sequenza di geni responsabili delle stesse caratteristiche ereditarie.
Ad esempio, se un gene che influisce sul colore dei petali è situato in un determinato punto (
locus; plurale loci) di un cromosoma, allora il cromosoma omologo possiede un gene simile per il colore dei petali nello stesso punto. Contrariamente a quanto avviene nel caso dei cromatidi fratelli, che sono effettivamente identici, in quanto derivati da duplicazione del DNA in interfase, i due loci corrispondenti sui due cromosomi omologhi possono costituire versioni leggermente differenti. (ad es. può esservi un gene per la formazione di petali rosa su un cromosoma e un gene per l'assenza di petali rosa nel medesimo locus sul cromosoma omologo; per stabilire quale prevarrà ne parleremo a proposito di dominanza e recessività dei caratteri genici).
Questo schema si riferisce ad una sola coppia di cromosomi omologhi.
 1)
1) Ogni cromosoma si origina in seguito alla duplicazione della cromatina durante l'interfase che precede la meiosi.
2) La prima divisione, meiosi 1, porta alla segregazione dei due cromosomi della coppia di omologhi, distribuendoli a due cellule figlie distinte (aploidi).
In questa fase ogni cromosoma è ancora costituito da due cromatidi.
3) Durante la meiosi 2 i due cromatidi fratelli si separano e ciascuna delle quattro cellule figlie è aploide e contiene un singolo cromosoma della coppia di omologhi.
3.6 L'origine della variabilità geneticaCome abbiamo già detto, lo zigote che deriva dalla riproduzione sessuata risulta diverso geneticamente dalle due piante genitrici che l'hanno generato. In seguito vedremo che proprio tale variabilità genetica costituisce il materiale su cui agisce la selezione naturale. Per il momento vedremo invece in che modo si origina la variabilità genetica attraverso la meiosi e la fecondazione.
Assortimento indipendente dei cromosomiNella seguente figura viene illustrato uno dei modi in cui la meiosi contribuisce alla variabilità genetica. La figura mostra come la disposizione delle coppie di cromosomi omologhi durante la metafase 1 della meiosi influisce sui gametofiti prodotti. Anche in questo caso l'esempio di riferisce ad un organismo diploide provvisto di quattro cromosomi omologhi (2 coppie), dove i colori rosso e blu servono per evidenziare i cromosomi omologhi. Questi colori sottolineano il fatto che i cromosomi omologhi differiscono geneticamente, sebbene quando osservati al microscopio ottico essi morfologicamente si somiglino notevolmente. Ad esempio, il cromosoma 3 ereditato da una delle piante genitrici porta indubbiamente molti geni, leggermente differenti da quelli situati sul cromosoma 3, ereditato dall'altra pianta genitrice.


L'orientamento delle varie coppie di omologhi (ogni coppia di omologhi viene anche definita tetrade, in quanto formata da quattro cromatidi, due appartenenti a un omologo e due all'altro) durante la metafase 1 risulta del tutto casuale e pertanto nel nostro esempio esistono due possibili modi di allineamento dei cromosomi durante la metafasi 1. Nel primo caso, le tetradi sono orientate con entrambi i cromosomi rossi su un lato. In questo caso, ognuno dei quattro gameti prodotti alla fine della meiosi 2 possiede soltanto cromosomi rossi o soltanto cromosomi blu (combinazioni 1 e 2). Nel secondo caso le tetradi sono orientate diversamente e ciò porta alla produzione di gameti contenenti un cromosoma rosso e uno blu. Inoltre, metà dei gameti possiede un grande cromosoma blu e un piccolo cromosoma rosso (combinazione 3), mentre l'altra metà possiede un grande cromosoma rosso e un piccolo cromosoma blu (combinazione 4).
Risulta pertanto evidente che in questo esempio i gameti prodotti possono avere quattro possibili combinazioni cromosomiche e l'organismo produrrà infatti gameti dei quattro diversi tipi. Questa varietà nei gameti si origina poiché durante la metafase 1 ogni coppia di cromosomi omologhi si orienta indipendentemente dall'altra coppia di omologhi. In una specie provvista di un maggiore numero di coppie cromosomiche, come ad esempio un'orchidea, ogni coppia di cromosomi si orienta indipendentemente dalle altre durante la metafase 1. In ogni specie, il numero totale di combinazioni cromosomiche possibili è 2
n, dove n corrisponde al numero aploide. Per quanto riguarda l'organismo a cui fa riferimento la figura, n=2, cossichè il numero di combinazioni cromosomiche è 2
2 = 4. Nella nostra specie (n=23) ci sono 2
23, ovvero circa 8 milioni di possibili combinazioni cromosomiche! Ciò significa che ogni gamete prodotto contiene una fra circa 8 milioni di possibili combinazioni di cromosomi materni e paterni.
Fecondazione casualeQuante sono le possibilità che un certo gamete di un individuo si unisca proprio con un determinato gamete di un altro individuo durante la fecondazione? Nella nostra specie, una cellula uovo, che contiene una fra circa 8 milioni di possibili combinazioni cromosomiche, viene fecondata a caso da uno degli spermatozoi che contiene una di circa 8 milioni di altre possibili combinazioni cromosomiche. E lo stesso vale per i gametofiti delle orchidee. Moltiplicando 8 milioni per 8 milioni, ne deriva che un uomo e una donna possono produrre uno zigote diploide provvisto di una fra 64.000 miliardi di possibili combinazioni cromosomiche! Se riprendiamo l'esempio della
Orchis purpurea (2n= 42; n= 21) ecco che il gametofita prodotto contiene una fra circa 2 milioni di possibili combinazioni di cromosomi delle due piante parentali (ovviamente tutto questo discorso vale solo nel caso della impollinazione incrociata tra due piante; nel caso di autoimpollinazione o geitonogamia il discorso varia!) e la futura pianta nascitura sarà una fra 4.400 miliardi di possibili combinazioni! E' dunque evidente che la modalità casuale della fecondazione incrociata aggiunge una quantità enorme di potenziale variabilità agli esemplari prodotti in seguito alla riproduzione sessuata. Questi numeri estremamente alti suggeriscono che l'orientamento indipendente dei cromosomi durante la metafase 1 e la fecondazione casuale potrebbero da soli essere responsabili di tutta la variabilità che si può osservare tra gli esemplari di una stessa specie. In realtà, questi due fattori costituiscono soltanto una parte del quadro.
Crossing overFinora è stata descritta la variabità genetica dei gameti e dello zigote a livello di interi cromosomi, ma abbiamo tralasciato un importante fenomeno, chiamato crossing over, ovvero lo scambio di frammenti cromosomici corrispondenti tra due cromosomi omologhi. Tale processo si verifica durante la profase 1 della meiosi. Nella figura è illustrato il crossing over tra due cromosomi omologhi e la successiva formazione dei gameti. Al momento dello scambio di materiale genetico, i cromosomi omologhi sono strettamente accostati lungo il loro asse maggiore secondo un preciso allineamento gene-gene. Al microscopio ottico i siti di crossing over sono visibili sotto forma di regioni a ×, definite
chiasmi (dal greco "incrocio"). In corrispondenza dei chiasmi i cromosomi omologhi rimangono uniti fino all'anafase 1.
Lo scambio di segmenti tra cromosomi omologhi (non tra cromatidi fratelli di uno stesso cromosoma in quanto geneticamente identici) aumenta la variabilità genetica che dervia dall'orientamento indipendente delle coppie di cromosomi durante la metafase 1. In assenza di crossing over, nell'esempio riportato nella figura, si arriverebbe alla produzione di due soli tipi di gameti, ovvero si avrebbero soltanto cromosomi parentali rossi o blu (come avevamo visto nell'immagine dell'Assortimento indipendente dei cromosomi). Il fenomeno dei crossing over permette la produzione di gameti provvisti di cromosomi in parte rossi e in parte blu. Questi cromosomi vengono definiti "ricombinanti", poiché essi derivano dalla
ricombinazione genetica, ovvero dalla produzione di combinazioni geniche diverse da quelle presenti nei cromosomi parentali. Dal momento che la maggior parte dei cromosomi contiene migliaia di geni, un singolo evento di crossing over può coinvolgere molti geni. Se si considera inoltre che in ogni tetrade possono verificarsi contemporaneamente numerosi crossing over, non sorprende che i gameti e la prole mostrino di regola una notevole variabilità.
Finora sono state esaminate tre fonti di variabilità genetica negli organismi a riproduzione sessuata: Crossing over durante la profase 1 della meiosi, Orientamento indipendente delle coppie cromosomiche durante la metafase 1 e la fecondazione casuale. In seguito vedremo una ulteriore fonte di variabilità genetica, legata alla genetica molecolare, ovvero le mutazioni, che costituiscono un evento piuttosto raro e che generano cambiamenti nel DNA dei geni. Le diverse versioni geniche che si trovano sui cromosomi omologhi si sono inizialmente originate proprio grazie a eventi mutazionali e, in ultima analisi, le mutazioni sono responsabili della diversità genetica che caratterizza gli organismi viventi.